We use cookies to make your experience better. To comply with the new e-Privacy directive, we need to ask for your consent to set the cookies Learn more.
AUTOMATED ACCURACY
BMDS™ provides advanced state-of-the-art technology for the safest, most accurate identification of research models available today, creating fully integrated systems designed to automate gathering, recording, and validating data for researchers and facilities. BMDS™ is the longest-running RFID solution for laboratory research, with industry-proven, dependable products and impeccable service.
BMDS™ – Bio Medic Data Systems™
Call (800) 526-2637 or (302) 628-4100
819 Bakke Avenue
Waterford, Wisconsin 53185 USA
Via Email
For Order Related inquires such as a request for quote or purchase orders, please use bmds.orders@avidityscience.com
For Technical Support and questions regarding our products, please use bmds.productsupport@avidityscience.com
For Payments and Remittances, please use bmds.ar@avidityscience.com
|
For Orders, POs or |
For general inquiries, technical support or product questions: |
For payments and |
 |
|
|
BENEFITS TO UPGRADE TO MICROCHIP / RFID TECHNOLOGY
|
|
|
|
|
|
Accuracy |
Efficiency |
Consistency |
Safety |
|
Fail-Safe Data Integrity Remove the manual
|
Save Time with Digital Data Easy portable readers allow |
Safe, Single Permanent Other applications are short-term and require continuous maintenance. Insertion of RFID transponder is easy with one integrated implant system. Transponders are coated with anti-migration technology to keep your animals safe and properly |
Improves Animal Welfare with Repeat ID applications can cause harm and stress to the animals. Avoid excessively handling your animals by inserting RFID technology at the beginning of your research. |
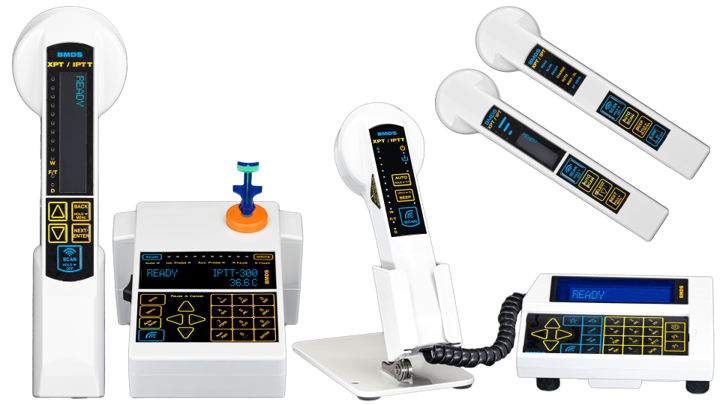
FEATURES OF THE BMDS™ PORTFOLIO
BMDS™ RFID technology is designed specifically for animal research with IACUC / AAALAC standards in mind.
Great service offering that backs durability and quality of product. Supports product through upgrades and servicing.
Sole programmable / temperature animal transponder made in the USA.
Only animal transponder that has been validated with a 2-year chronic evaulation for safe and effective us in all animal models.
|
BMDS™ PORTFOLIO OVERVIEW |
||||
|
|
Read-Only | Programmable | Programmable with Temperature | |
|
|
Cordless | Stationary | Multiple Interface Options | |
Compatible with all Laboratory Information Management Systems (LIMS) including:



 United Kingdom
United Kingdom